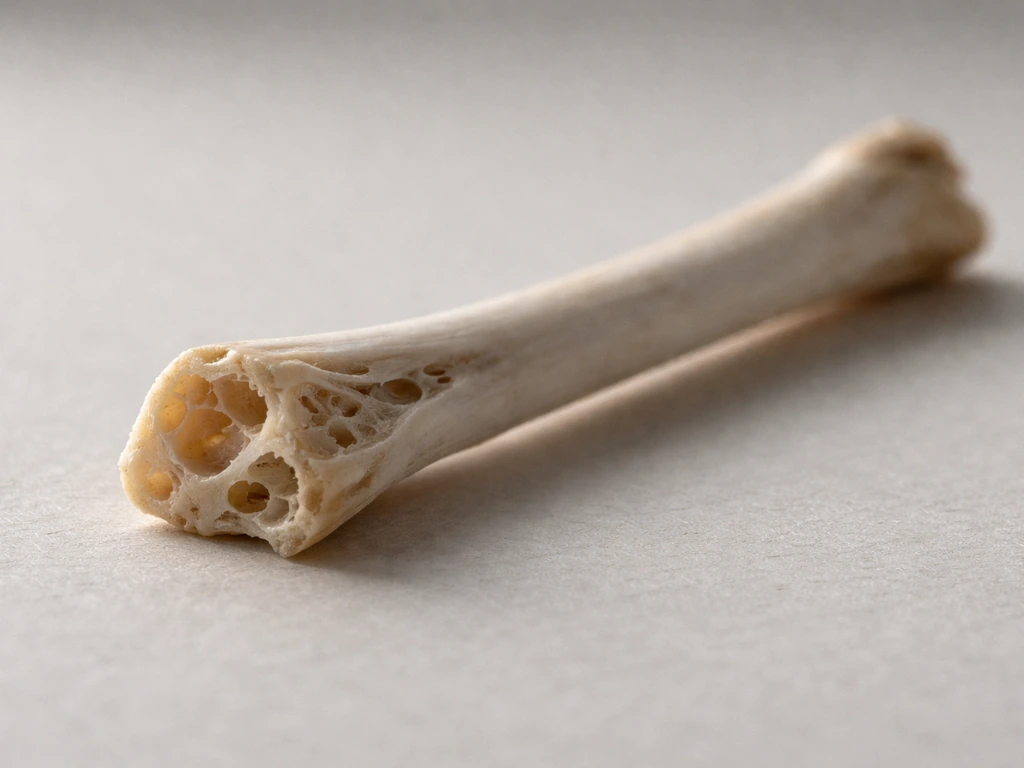
Bird bones are made of the same fundamental stuff as your own bones: a tough protein mesh called collagen woven together with a hard mineral called hydroxyapatite, plus a small amount of water. What makes bird bones genuinely different from mammal bones is not the raw materials but the architecture built from those materials. Birds have evolved thinner cortical walls, air-filled hollow spaces connected to the respiratory system, and special temporary bone tissue for calcium storage during egg-laying. Together, these features produce a skeleton that can be 13–25% lighter than a comparable mammal skeleton while still handling the enormous forces of flapping flight.
What Are Bird Bones Made Of: Composition, Strength & Function

Marcus Chen
15 Jul 2026
Why bird bone composition is worth understanding
I got curious about this after holding a pigeon wing bone at a natural history museum and being genuinely surprised by how light it felt. It almost seemed like a trick. That sent me down a rabbit hole into avian skeletal biology, and what I found was that the chemistry and structure of bird bones are actually a masterclass in biological engineering. If you are a student trying to understand vertebrate biology, a bird enthusiast who has ever wondered why a hawk can pull off aerial maneuvers that would snap a human arm, or someone who just found a bone in the garden and wants to know what it is made of, the composition of bird bones gives you the answer to a surprisingly large number of follow-up questions, including how strong bird bones really are, whether they break easily, why bird fossils are relatively rare, and how birds connect evolutionarily to dinosaurs.
The main components of bird bones at a glance
Bird bone tissue is a nanocomposite, which just means it is made of two distinct materials blended at a very fine scale. One component is hard and mineral-based; the other is soft and protein-based. Neither alone would do the job well. The mineral without the protein would be brittle like chalk. The protein without the mineral would be flexible but far too weak to bear load. Together they achieve a balance of stiffness, strength, and resistance to fracture that no single material can match.
| Component | Approximate proportion by weight | Role in bone |
|---|---|---|
| Mineral (hydroxyapatite) | 60–70% | Provides compressive strength and stiffness |
| Organic matrix (mainly collagen) | 20–30% | Provides tensile strength, flexibility, and toughness |
| Water | 5–10% | Maintains tissue hydration and contributes to mechanical behavior |
These proportions are consistent across vertebrates as a group, but they shift with age, the specific bone region, the bird species, and measurement method. A highly pneumatized humerus from an albatross and a dense leg bone from an ostrich are both made of collagen and hydroxyapatite, but the ratios, thicknesses, and internal geometry are dramatically different.
Collagen: the flexible protein framework inside every bone
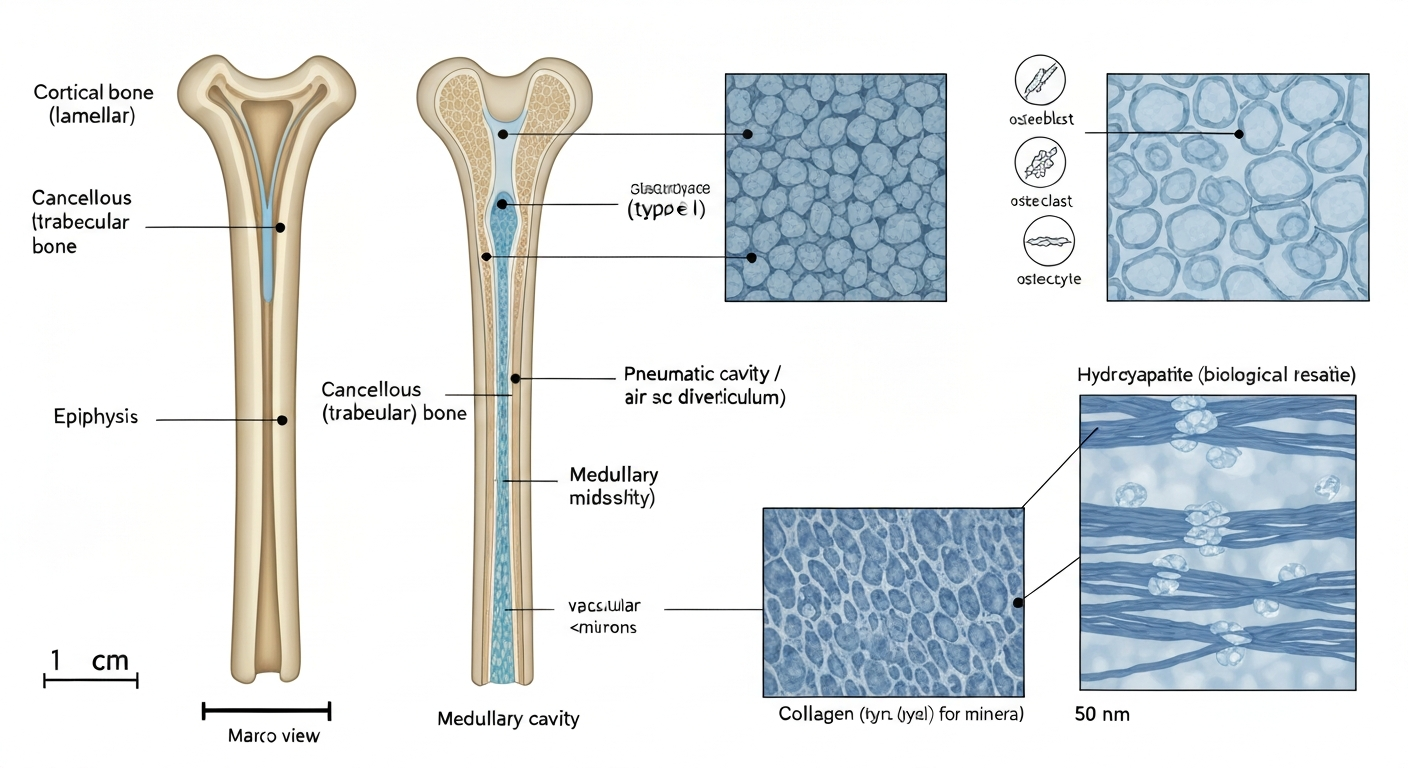
The organic phase of bone is mostly type I collagen, which is the same structural protein found in tendons, skin, and ligaments. Collagen molecules self-assemble into stiff fibers that can resist being pulled apart (tensile loading) without snapping. In bone, these fibers are laid down by cells called osteoblasts and then mineralized, which is where the hydroxyapatite crystals come in. Think of the collagen as reinforcing rods and the mineral as the concrete poured around them: neither is great alone, but the combination resists both compression and tension.
The orientation of collagen fibers within a layer of bone also matters a great deal for mechanical performance. In rapidly grown bone (called woven bone), fibers are arranged somewhat randomly, giving moderate strength in multiple directions. In slower, more mature bone, fibers are organized in orderly sheets called lamellae (each sheet is a lamella), producing a tissue called lamellar bone that is significantly stiffer and stronger. Most of the load-bearing cortical bone in adult birds is lamellar. The ratio of mineral to collagen is one of the primary controls on whether bone is stiff and somewhat brittle (more mineral) or flexible and tougher (more collagen). Higher mineral content pushes bone toward compressive strength at the cost of reduced ability to absorb energy before fracturing.
Hydroxyapatite: how bone gets its hardness
The mineral in bird bone is not pure hydroxyapatite as you would find in a chemistry lab. It is a biological version that is laced with small amounts of carbonate, fluoride, magnesium, and other ions substituted into the crystal lattice. This ion-substituted biological apatite forms tiny, plate-shaped crystals just nanometers in size that nestle between and within collagen fibrils. The result is a mineralized composite tissue that has a measured flexural stiffness (Young's modulus, which describes how much a material bends under load) ranging from roughly 9 to 21 GPa in avian long bones, with values shifting depending on the species, the specific bone, and how pneumatized it is. Distal wing bones like the radius and the tibiotarsus tend to show higher stiffness values than the humerus or femur in comparative datasets. Mechanical tests compiled in Avian long bones, flight and bipedalism (Cubo & Casinos 2001) show that bone material properties and whole-bone strength scale with size and vary by element, with distal wing elements (radius) and tibiotarsus often exhibiting higher mean bending strength and Young's modulus than humerus or femur Avian long bones, flight and bipedalism (Cubo & Casinos 2001) — compiled mechanical data.
Mineralization is not a one-time event at birth. It is an ongoing process regulated throughout a bird's life. The skeleton is continuously broken down and rebuilt, with cells removing old mineral and other cells depositing new crystal structures. This is how birds manage calcium demands for egg production, how bones repair after injury, and how the skeleton adapts to changing loads over a bird's lifetime.
The cells that build, maintain, and remodel bird bone
Bone is a living tissue, which still surprises some people who assume it is more like a rock than an organ. Three cell types do most of the work in bird bone, just as they do in all vertebrate bone.
- Osteoblasts are the bone-builders. They secrete the collagen matrix and trigger its mineralization, essentially constructing new bone tissue layer by layer.
- Osteoclasts are the bone-dissolvers. They break down existing bone matrix and release calcium back into the bloodstream, which is essential for remodeling and for mobilizing calcium reserves when demand is high.
- Osteocytes are mature osteoblasts that have become embedded in the bone matrix they built. They sit inside tiny spaces called lacunae and communicate through a network of fine channels, acting as mechanical sensors that signal when bone needs to be strengthened or remodeled.
At the histological (microscopic tissue) level, avian cortical bone can contain secondary osteons, also called Haversian systems. An osteon is a roughly cylindrical unit of bone built concentrically around a central blood vessel channel, with lamellar bone rings surrounding it. Birds vary considerably in how much Haversian remodeling they show, with some species and bones showing dense osteon populations and others remaining largely in a primary, unmodeled state. One particularly interesting microstructural adaptation in bird wings is a high proportion of laminar bone, primary cortical bone in which the vascular canals run in circular rings around the bone's circumference rather than lengthwise. Laminar bone as an adaptation to torsional loads in flapping flight (de Margerie 2002, J Anat) documented high laminarity concentrated in the humerus, ulna, femur and tibiotarsus across multiple bird species, interpreting this circular vascular arrangement as a microstructural response to habitual twisting during flapping. Research by Emmanuel de Margerie and colleagues published in 2002 identified this laminar arrangement in the humeri, ulnae, femora, and tibiotarsi of multiple species and interpreted it as an adaptation to resist the torsional (twisting) loads generated by flapping. It is the kind of microstructural detail you would only see under a microscope, but it has real mechanical consequences for how a wing bone handles the stress of each wingbeat.
Cortical bone versus cancellous bone: two structures, two jobs
Pick up any bird skeleton and you are mostly looking at cortical bone, also called compact bone. It forms the dense outer shell of every bone in the body and is responsible for the majority of load-bearing strength. In birds, cortical walls tend to be notably thinner than in mammals of comparable body mass, which is how the skeleton saves weight. But thinner walls alone would make bones weaker, so birds compensate using geometry: placing a thin, stiff tube at a large radius (wide bone diameter) is actually a very efficient way to resist bending and torsion. The second moment of area, which describes how spread-out the bone material is from the central axis, increases with the square of the radius, so a wider tube with thin walls can outperform a narrower tube with thick walls at a fraction of the weight. This is exactly the principle behind hollow steel tubes in engineering.
Cancellous bone, also called spongy or trabecular bone, is the lattice-like tissue found inside bone ends (epiphyses) and in the interior of some elements. It looks like a three-dimensional mesh or a honeycomb under a microscope. In birds, internal trabecular reinforcement is especially important inside pneumatized bones, where struts and ridges of cancellous bone resist the tendency of a thin-walled hollow tube to buckle or collapse under load. The ends of long bones, where complex joint forces meet, also rely heavily on this spongy interior architecture to distribute stress over a wider area and prevent local failure.
| Bone type | Structure | Location in skeleton | Primary mechanical role |
|---|---|---|---|
| Cortical (compact) | Dense, solid outer shell with lamellar organization | Shaft of all long bones, outer layer of flat bones | Resists bending, torsion, and compressive loads |
| Cancellous (spongy/trabecular) | Open lattice of interconnecting bone struts (trabeculae) | Bone ends (epiphyses), interior of pneumatized bones | Distributes joint loads, resists buckling, reduces weight |
| Medullary bone | Woven, highly mineralized, porous temporary tissue | Marrow cavities of many skeletal elements in laying females | Rapid calcium reserve for eggshell production |
Pneumatic bones: when air is part of the skeleton
One of the most genuinely fascinating things I learned is that many bird bones are not just hollow in the everyday sense but are actually extensions of the respiratory system. Diverticula, which are small outpouching sacs connected to the air sac system, invade the bone interior during development and replace the marrow with air. These are called pneumatic bones. In a large soaring bird like a pelican or frigate bird, a substantial proportion of the skeleton is pneumatized, including the humerus, vertebrae, sternum, and sometimes bones of the pelvis and skull.
A 2023 microCT study that scanned humeri across a broad range of bird species directly quantified the air fraction (pneumaticity score) in each specimen and found that pneumaticity is taxonomically widespread but patchy, meaning closely related species can differ considerably in how much of their humerus is air. This kind of quantitative data is now archived in open repositories like MorphoSource, which is making it possible to compare internal bone architecture across hundreds of species without destroying any specimens.
Mechanically, pneumatized bones have thinner cortical walls and lower measured bending strength per unit cross-sectional area than non-pneumatic bones of similar outer dimensions. The trade-off is reduced mass. Birds compensate for the structural thinning by maintaining internal trabecular struts that brace the bone wall from within, and by relying on circular cross-sectional geometry that efficiently resists the twisting and bending that flapping imposes. It is worth noting that thin cortical walls and high pneumaticity do make bones more vulnerable to catastrophic fracture from sharp localized impacts, a point that matters practically when thinking about bird injuries. Whether bird bones break easily is partly a function of this architecture alongside the loads involved.
Which bones are typically pneumatized
- Humerus (upper wing bone): commonly pneumatized in many flying birds, especially larger species
- Vertebrae: cervical and thoracic vertebrae are pneumatized in many groups
- Sternum and ribs: often pneumatized in larger birds
- Skull bones: cavities in the skull of some species serve a similar weight-saving role
- Pelvis and synsacrum: pneumatized in some larger birds including many raptors
- Leg bones (femur, tibiotarsus): generally not pneumatized in most species, as they bear body weight during standing and walking
Medullary bone: a temporary calcium bank for egg-laying females
Medullary bone is something I had never heard of before diving into avian anatomy, and it is genuinely remarkable. Female birds in reproductive condition deposit a special type of bone tissue called medullary bone inside the marrow cavities of many skeletal elements, often two weeks before egg-laying begins. It is a woven, highly mineralized, and notably porous structure that is chemically and histologically distinct from the surrounding cortical bone. Its entire purpose is to act as a rapidly mobilizable calcium reserve: when the bird forms an eggshell, it needs enormous amounts of calcium very quickly, and medullary bone can be dissolved by osteoclasts and redeposited as eggshell mineral in a matter of hours.
The formation and resorption of medullary bone is controlled by estrogen. Research on house finches and pine siskins has confirmed that higher circulating estrogen levels correlate with greater medullary bone quantity and density. Experimentally raising estrogen in birds prompts medullary bone deposition even outside the normal breeding season. After egg-laying ends, the tissue is typically resorbed and the marrow cavity returns to its usual state.
Medullary bone forms systemically, meaning it appears in many bones at once, not just in one convenient reservoir. It has been identified in the femur, tibiotarsus, humerus, ulna, ribs, and even some smaller elements. This wide distribution makes sense because the bird needs to mobilize calcium rapidly and from multiple sites simultaneously. Paleontologists have also identified medullary bone in fossil specimens, including some non-avian dinosaur fossils, which has been used as evidence of reproductive state in extinct species, though this area is still debated and subject to ongoing research. It is a reminder that the tissues in bird bones are not just mechanically important but physiologically active in ways that connect to behavior, reproduction, and evolutionary history.
Fused and specialized skeletal elements unique to birds
Beyond tissue composition, the bird skeleton has several distinctive fused or modified elements that are worth mentioning because they come up constantly in discussions of avian anatomy. The furcula, or wishbone, is two clavicles fused into a single springy U-shaped structure that stores and releases elastic energy during the downstroke of the wing. The synsacrum is a fusion of several lumbar, sacral, and caudal vertebrae into a single rigid unit that connects the pelvis and hind limbs to the spine, providing a stable platform for bipedal walking and perching. The pygostyle, at the tail end, is several fused caudal vertebrae that anchor the tail feathers. All of these fused elements are made of the same collagen-hydroxyapatite composite tissue as any other bone; they are architectural innovations, not chemical ones.
Bird bones versus dinosaur bones: more similar than you might expect
The connection between bird bones and dinosaur bones is not just a phylogenetic footnote. At the histological level, the bones of many non-avian dinosaurs share features with birds that are clearly ancestral: fibrolamellar bone tissue (a woven-to-lamellar mix associated with fast growth), high vascularization, and in some theropod dinosaurs, evidence of medullary bone and pneumaticity. Reviews of dinosaur paleohistology by researchers building on the work of Armand de Ricqlès, Jack Horner, and Kevin Padian have traced these shared features through the dinosaur-to-bird transition and shown that birds effectively inherited a skeleton already predisposed toward fast growth and active metabolism.
Where birds diverge from most non-avian dinosaurs in bone histology is in the degree of cortical thinning, the systematic development of extensive pneumaticity throughout the skeleton, and the appearance of medullary bone tied to a rapid egg-laying reproductive strategy rather than the slower egg-production cycles typical of large theropods. So a bird femur and a theropod dinosaur femur are both built from the same cellular machinery and the same mineral-collagen composite, but birds have taken the architecture to an extreme that non-avian dinosaurs never quite reached.
How composition affects strength, fragility, and fracture risk
A question I see come up constantly is whether bird bones are weaker than mammal bones. For a concise overview of comparative strength and typical measurements, see the related article how strong are bird bones. For a focused discussion on whether bird bones break easily, see do bird bones break easily. The honest answer is: it depends what you mean by weak. At the material level (comparing a cubic millimeter of bird cortical bone to a cubic millimeter of mammal cortical bone), the tissues are broadly comparable in stiffness and strength. The differences emerge at the structural level. Thin cortical walls and air-filled interiors reduce the total cross-sectional material resisting a load, which means a given external force creates higher stress in the bone wall. This does make pneumatized bones more vulnerable to catastrophic fracture when struck by a concentrated local force, like a sharp impact or a collision.
Birds manage this vulnerability through geometry (wide circular cross-sections, internal struts, ridges and reinforcements at high-stress regions) and through behavioral and postural strategies that distribute loads broadly across the skeletal architecture rather than concentrating them. A wing bone during flight is loaded in a complex combination of bending and torsion spread across its length rather than struck at one point. The skeleton is designed for those distributed, predictable loads, not for unpredictable point impacts from collisions, which is precisely why window strikes and handling injuries can fracture even robust-looking bird bones.
Why bird bones are harder to find as fossils
If bird bones are made of the same mineralized tissue as dinosaur bones, why are bird fossils so much rarer? The answer lies in taphonomy, the science of how organisms become (or fail to become) fossils. Studies that have quantified preservation biases in the avian fossil record have shown that small, thin-walled, highly pneumatized bones are predictably under-represented compared to larger, denser bones. They fragment more easily after death, are consumed and scattered by scavengers, and decompose or are crushed before burial can protect them. Even when a bird dies near sediment suitable for preservation, delicate hollow bones often disintegrate before fossilization begins.
This does not mean bird fossils are impossible to find. For a focused discussion on whether bird bones fossilize, see the article Do bird bones fossilize. Some depositional environments, particularly fine-grained lake sediments and certain volcanic ash deposits, preserve delicate bird remains in remarkable detail. But the baseline expectation should be that avian bone composition and architecture make birds less likely to enter the fossil record intact compared to larger, more heavily built animals. It is a direct consequence of the same architectural choices that allow birds to fly.
How it all fits together: composition, structure, and flight
Once you understand what bird bones are made of and how they are arranged, the whole picture of avian flight mechanics starts to make more sense. The collagen-hydroxyapatite composite provides a bone tissue that is simultaneously stiff enough to transmit forces and tough enough not to shatter under cyclic loading. The thin cortical walls and pneumatized interiors reduce mass without sacrificing bending stiffness, because wide-diameter tubes are geometrically efficient at resisting bending even with minimal wall thickness. Laminar microstructure in wing bones specifically resists the torsional loads of flapping. Medullary bone shows that the skeleton is not just mechanical infrastructure but a physiological organ that participates in reproductive biochemistry. And the fused elements like the furcula and synsacrum show how the same basic material can be shaped into energy-storing springs or rigid structural platforms depending on where in the body it is needed. For gamers wondering "is bird bones worth it Fallout 76", community guides and discussions evaluate the item's in-game value and tradeoffs.
The skeleton of a bird is not just a lighter version of a mammal skeleton. It is a thoroughly redesigned system that happens to use the same fundamental building blocks all vertebrates share. That realization, for me, was the satisfying payoff of the rabbit hole that started with holding one surprisingly light pigeon bone at a museum.
FAQ
What are bird bones made of at the chemical and material level?
Like other vertebrates, bird bone is a composite of an inorganic mineral phase and an organic phase. The mineral is a biological apatite (ion‑substituted hydroxyapatite) that provides stiffness and compressive strength; the organic phase is primarily type I collagen, which gives toughness and resistance to crack propagation. Bone also contains water and cells (osteoblasts, osteoclasts, osteocytes) embedded in the matrix. By mass, vertebrate bone is typically ~60–70% mineral, ~20–30% organic matrix and ~5–10% water, with variation by species, age and bone region.
How is avian bone organized histologically and cellularly?
Avian cortical bone contains the same principal cell types as other vertebrates: osteoblasts (make bone), osteoclasts (resorb bone) and osteocytes (mechanosensors). Microstructural patterns include primary woven/lamellar bone, regions of laminar bone (vascular canals oriented circularly to resist torsion), and in many taxa secondary osteons (Haversian remodeling). The amount and pattern of remodeling vary by species, element and age.
What special bone types do birds have that differ from other vertebrates?
Three key specializations: 1) Pneumatic bones — bones invaded by air sacs/diverticula and thus containing air spaces that reduce mass; 2) Medullary bone — a female, estrogen‑dependent, highly mineralized woven tissue deposited in marrow cavities during egg production as a labile calcium reservoir for eggshell formation; 3) Fused elements — skeletal elements fused for stability (e.g., synsacrum, furcula, pygostyle) that change load paths and reduce weight by eliminating joints.
How do these features make the avian skeleton lightweight yet strong enough for flight?
Birds combine material and geometric strategies. At the material level, a mineralized collagen composite gives a high stiffness‑to‑mass ratio. Architecturally, birds place thin, stiff cortical bone far from the neutral axis (high second moment of area) to maximize bending and torsional resistance per unit mass. Pneumaticity removes internal mass while sparing structural cortical walls, and laminar bone microstructure helps resist torsional loads from flapping. Fused elements simplify load transfer and reduce redundant mass. Together these adaptations deliver a high strength‑to‑weight skeleton optimized for flight.
Does having thin or pneumatic bones make birds fragile and likely to break bones easily?
Thin cortices and pneumaticity do increase vulnerability to catastrophic fracture under high local loads or impacts because there is less material to absorb energy. Birds mitigate this through geometric design (circular cross‑sections, internal trabeculae at joints, struts and ridges) and by distributing loads across the skeleton during flight and landing. Vulnerability also varies by species, element and behavior: small, highly pneumatic bones are more fragile than stout, marrow‑filled bones in large or terrestrial birds.
How strong is bird bone compared with other vertebrates?
Measured stiffness (Young’s modulus) of avian cortical bone typically lies in the range of ~9–21 GPa in compiled studies, with variation by element and species. Higher mineral content increases stiffness and compressive strength but lowers toughness. Whole‑bone bending strength and stiffness scale with size, flight style and internal architecture; some avian elements (e.g., radius, tibiotarsus) can show higher apparent bending resistance than the humerus or femur depending on species and pneumatization.
Next Article
Are Bird Bones Fragile? How They Stay Strong for Flight
Are bird bones fragile? Lightweight yet strong bones use air spaces and internal support to resist flight forces.